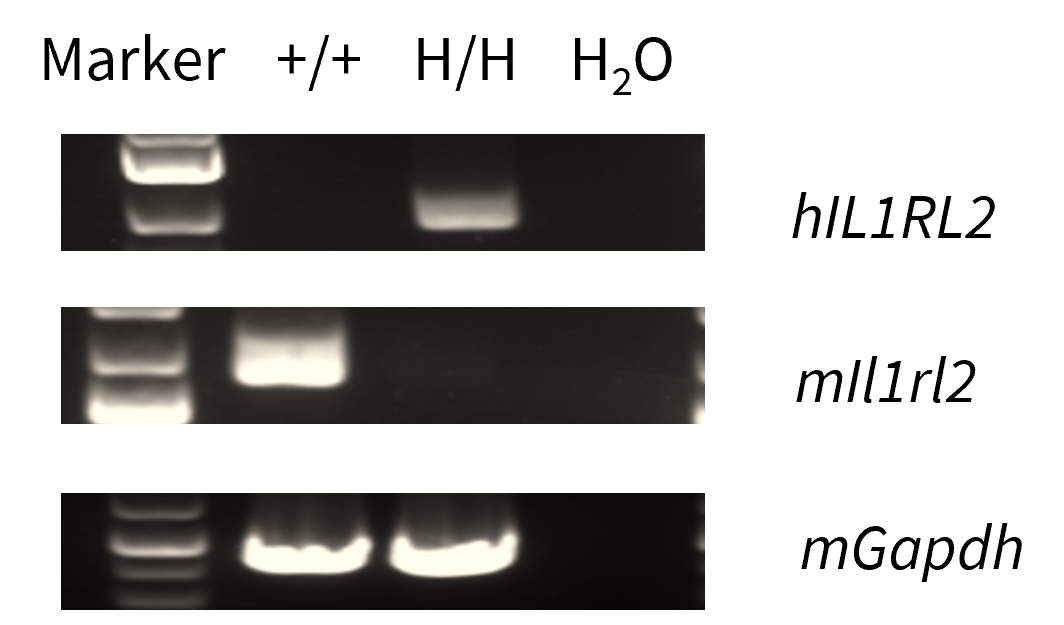
mRNA Expression Analysis in Lung
- Mouse Il36r mRNA was only detectable in wild-type mice.
- Human IL36R mRNA was exclusively detectable in homozygous B-hIL36R mice but not in wild-type mice.
Strain specific analysis of IL1RL2 mRNA expression in C57BL/6 mice and homozygous B-hIL36R mice by RT-PCR. Lung RNA was isolated from C57BL/6 mice (+/+) and homozygous B-hIL36R mice (H/H), then cDNA libraries were synthesized by reverse transcription, followed by PCR with mouse or human IL36R primers. Mouse Il36r mRNA was only detectable in wild-type mice. Human IL36R mRNA was exclusively detectable in homozygous B-hIL36R mice but not in wild-type mice.
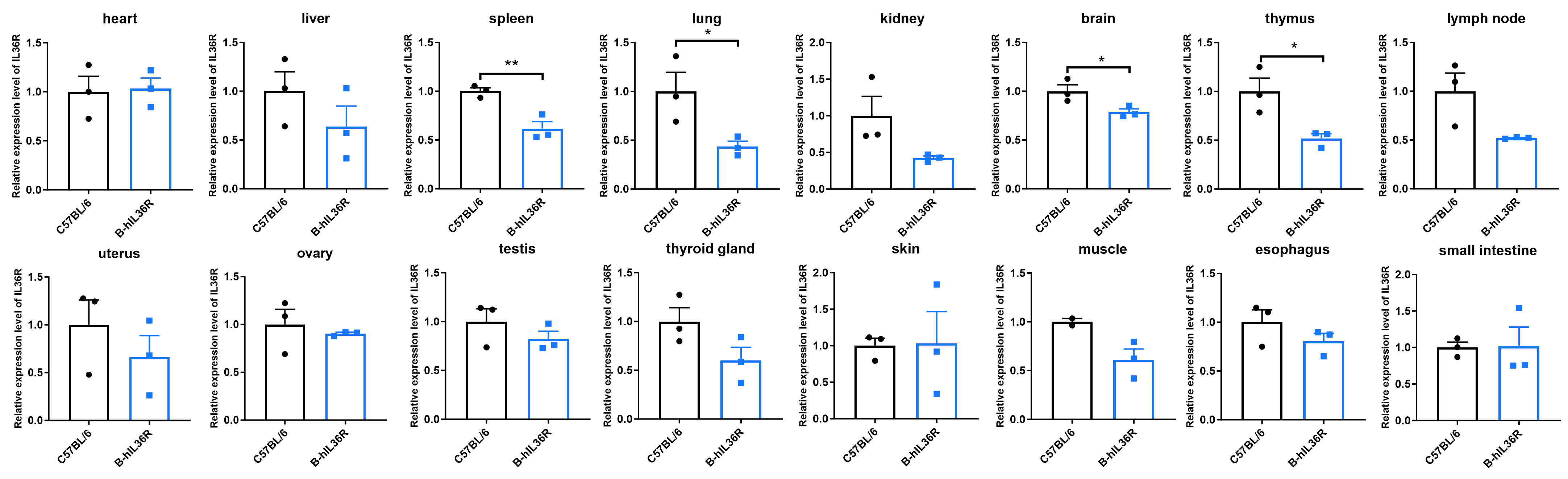
mRNA Expression Analysis
- The abundance of IL36R mRNA in spleen, lung, brain and thymus of B-hIL36R mice were significantly lower to those in WT mice, and the expression level of IL36R mRNA in other tissues of B-hIL36R mice were slightly lower or comparable than those in WT mice.
Quantitative PCR analysis of IL36R mRNA expression in wild-type (WT) mice and B-hIL36R mice. Total RNA was isolated from heart, liver, spleen, lung, kidney, brain, thymus, lymph node, uterus, ovary, testis, thyroid, skin, muscle, esophagus and small intestine of WT mice and homozygous B-hIL36R mice (n=3, 8 week-old), qPCR method was used to detect the RNA expression of IL36R. The results showed that the abundance of IL36R mRNA in spleen, lung, brain and thymus of B-hIL36R mice were significantly lower to those in WT mice, and the expression level of IL36R mRNA in other tissues of B-hIL36R mice were slightly lower or comparable than those in WT mice. The mean and standard deviation from triplicate samples are indicated.
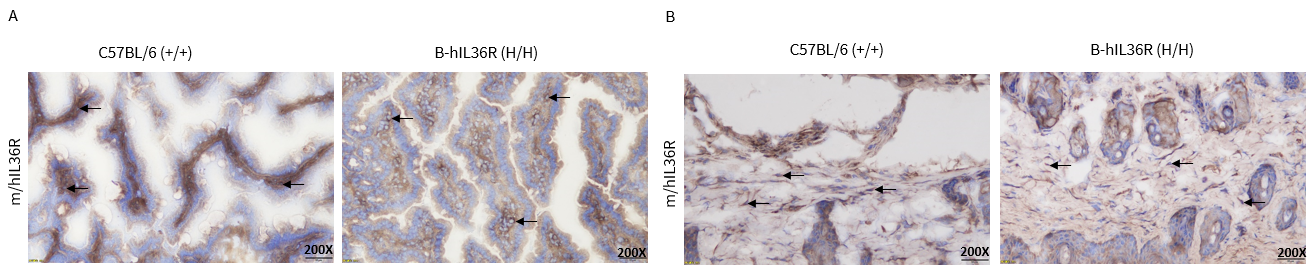
Protein Expression Analysis in Small Intestine and Skin
- IL36R was detectable in WT mice and homozygous B-hIL36R mice due to the cross-reactivity of antibodies.
Immunohistochemical (IHC) analysis of IL36R expression in homozygous B-hIL36R mice.
The small intestine and skin were collected from WT and homozygous B-hIL36R mice (H/H) that have established psoriasis models and prepared for OCT embedding. The tissues were analyzed by IHC with anti-IL36R antibody. (A), IL36R expression in the small intestine. (B), IL36R expression in the skin. IL36R was detectable in WT mice and homozygous B-hIL36R mice due to the cross-reactivity of antibodies. The arrow indicates tissue cells with positive IL36R staining (brown).
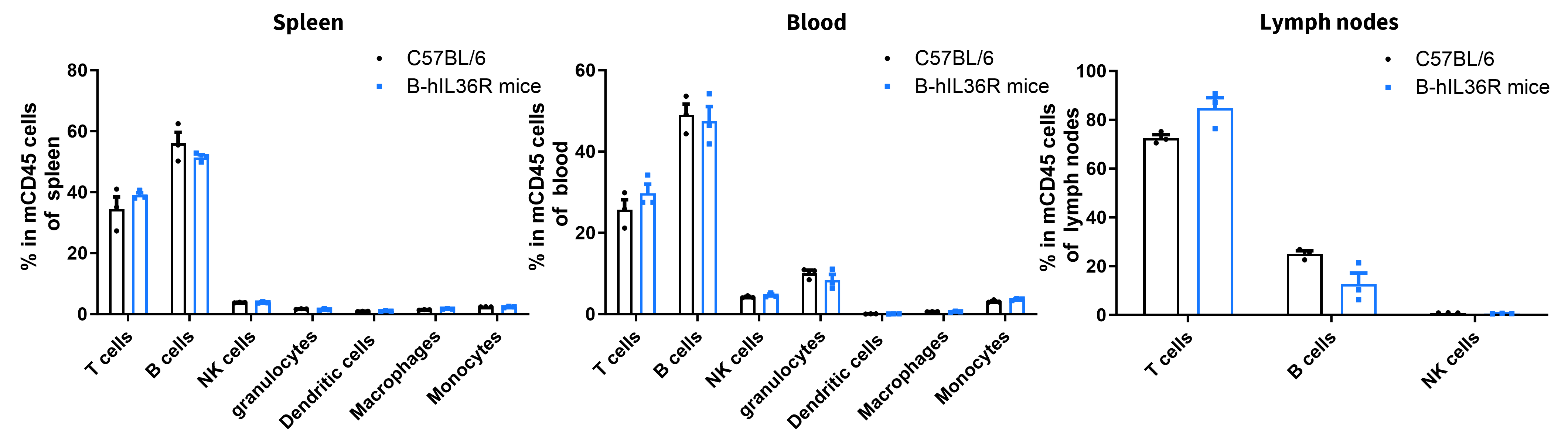
Analysis of Leukocyte Subpopulations
- Percent of T cells, B cells, NK cells, dendritic cells, granulocytes, monocytes and macrophages in homozygous B-hIL36R mice were similar to those in the C57BL/6 mice
- Humanization of IL36R does not affect normal immune cell development or splenic distribution.
Analysis of leukocyte subpopulations by FACS. Leukocytes were isolated from female C57BL/6 and B-hIL36R mice (n=3, 7 week-old). Flow cytometry analysis of the leukocytes was performed to assess leukocyte subpopulations. Percent of T cells, B cells, NK cells, dendritic cells, granulocytes, monocytes and macrophages in homozygous B-hIL36R mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hIL36R in place of its mouse counterpart does not change the overall development, differentiation or distribution of these cell types. Values are expressed as mean ± SEM.
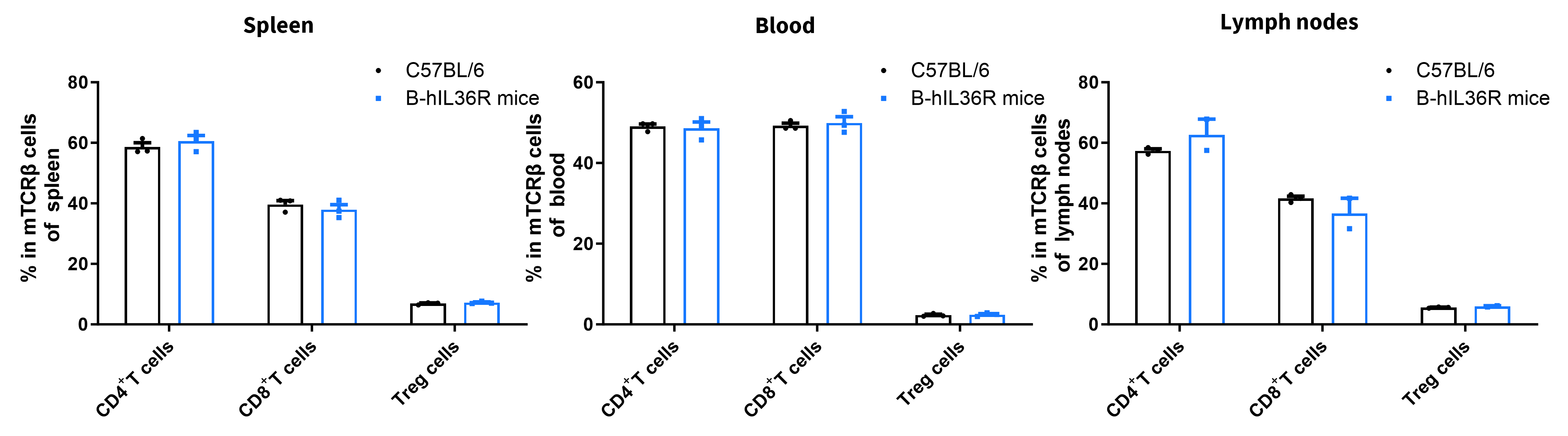
- Percent of CD8+ T cells, CD4+ T cells and Treg cells in homozygous B-hIL36R mice were similar to those in the C57BL/6 mice
- Humanization of IL36R does not change the overall development, differentiation or distribution of these T cell subtypes.
Analysis of T cell subpopulations by FACS. Splenocytes, peripheral blood and lymph nodes were isolated from female C57BL/6 and B-hIL36R mice (n=3, 7 week-old). Flow cytometry analysis of the splenocytes was performed to assess leukocyte subpopulations. Percent of CD8+ T cells, CD4+ T cells and Treg cells in homozygous B-hIL36R mice were similar to those in the C57BL/6 mice, demonstrating that introduction of hIL36R in place of its mouse counterpart does not change the overall development, differentiation or distribution of these T cell subtypes. Values are expressed as mean ± SEM.
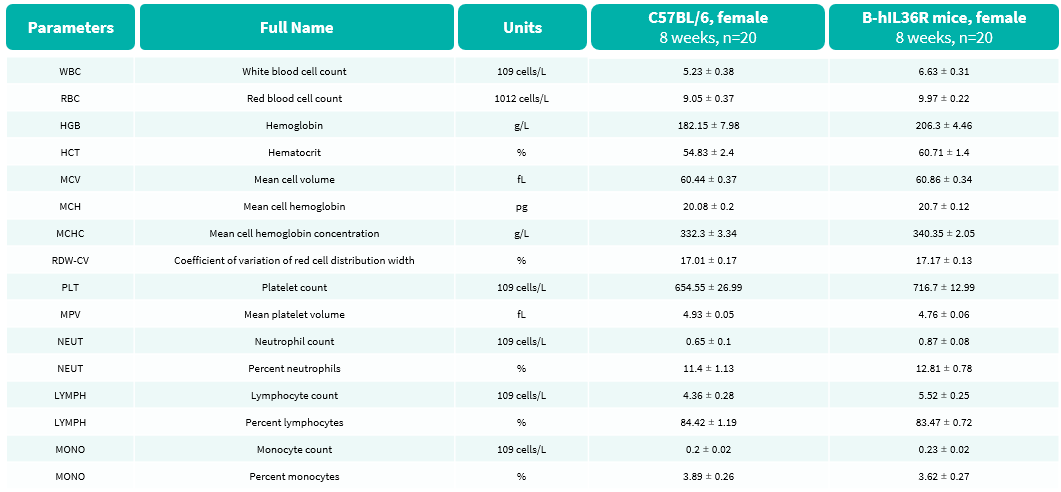
Hematology Analysis
- No significant differences were observed compared with wild-type mice.
Complete blood count (CBC) of B-hIL36R mice. Values are expressed as mean ± SD.
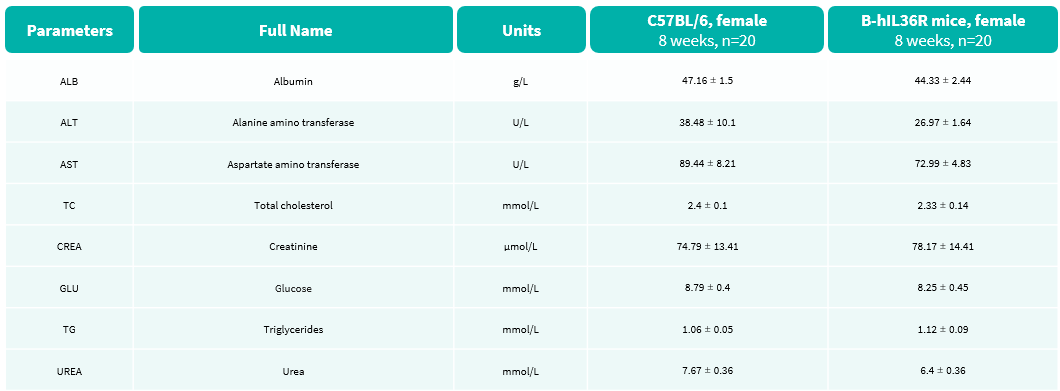
Blood Biochemical Analysis
- No significant differences were observed compared with wild-type mice.
Blood biochemical parameters of B-hIL36R mice are shown. Values are expressed as mean ± SD.
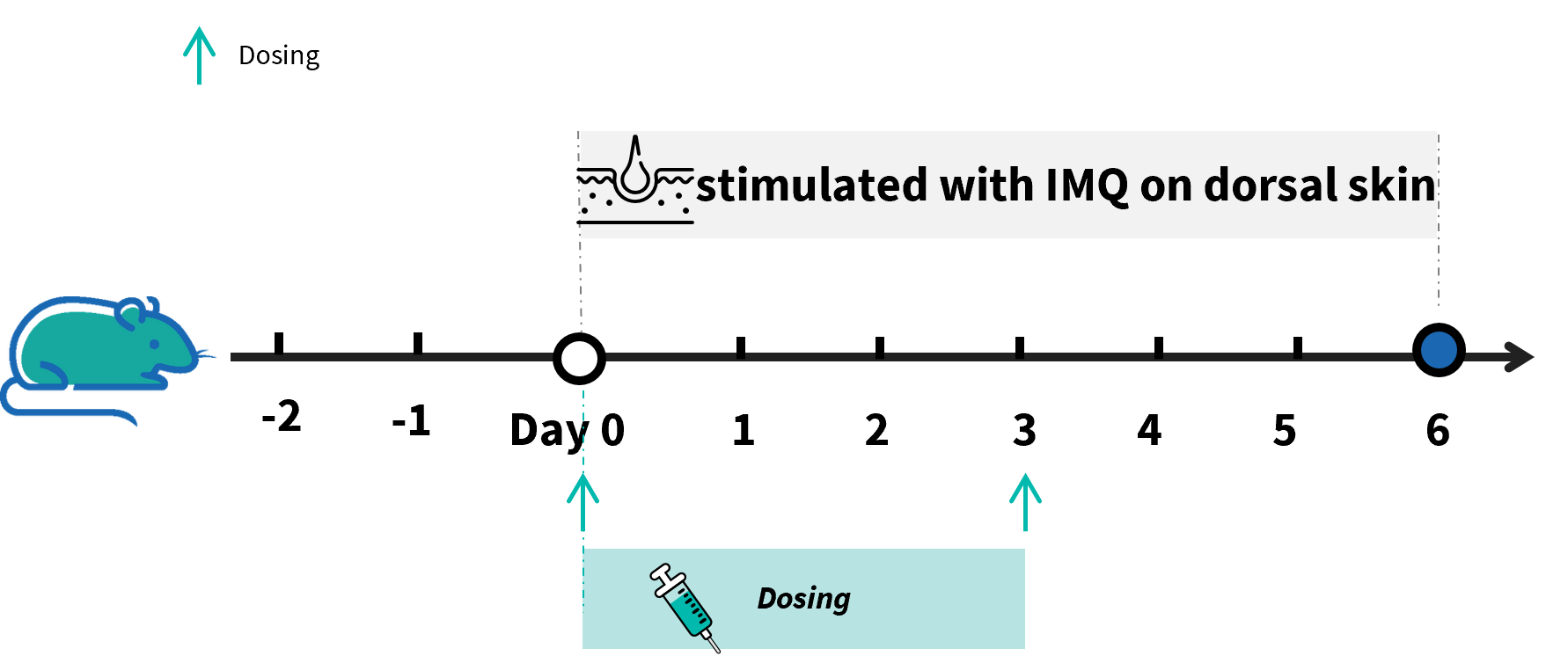
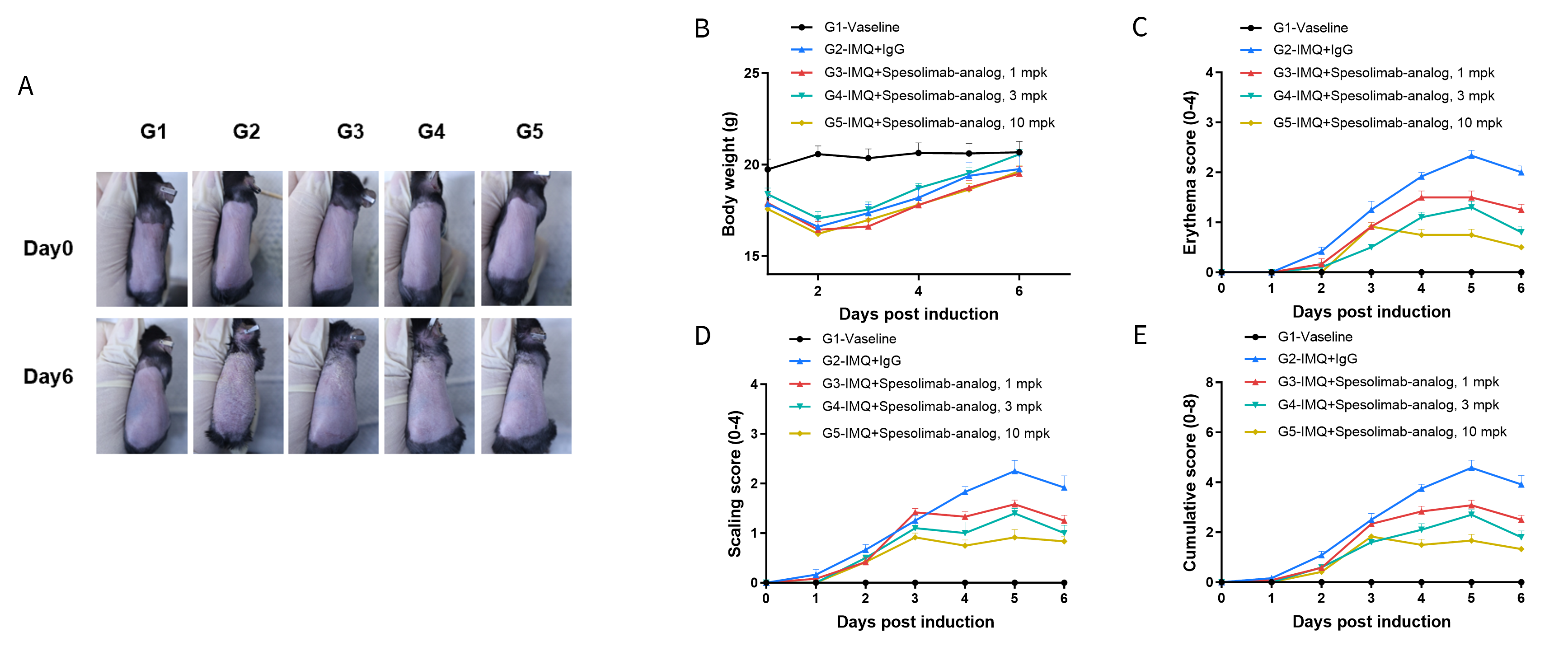
In Vivo Efficacy of Anti-human IL36R Antibodies in Psoriasis Model Induced in B-hIL36R Mice
Experimental schedule for IMQ-induced psoriasis model in B-hIL36R mice. Mice at 9 week-old of age received a daily topical of commercially available IMQ cream on the shaved back for 6 consecutive days to induce IMQ-induced psoriasis model. Control mice were treated similarly with Vaseline cream. Severity of skin inflammation was daily scored and back skin was collected at the endpoint. IMQ: imiquimod.
- After treatment of anti-hIL36R antibody, the phenotypical presentation level and erythema and scaling score of the back were much lower than the control in homozygous B-hIL36R mice.
IMQ-induced psoriasis model in B-hIL36R mice phenotypically resembles psoriasis. Mice (female, 9 week-old, n=6) were scored daily for up to 7 days for body weight and clinical signs of skin inflammation following treatment with imiquimod (IMQ) cream. Mice in each group were treated with different doses of Spesolimab-analog (in house). (A) Phenotypical presentation of mouse back skin at day 0 and day 6. (B) Body weight changes during treatment. (C-E) Erythema and scaling score of the back was scored daily. Additionally, the cumulative score (erythema plus scaling) is depicted. Values are expressed as mean ± SEM.
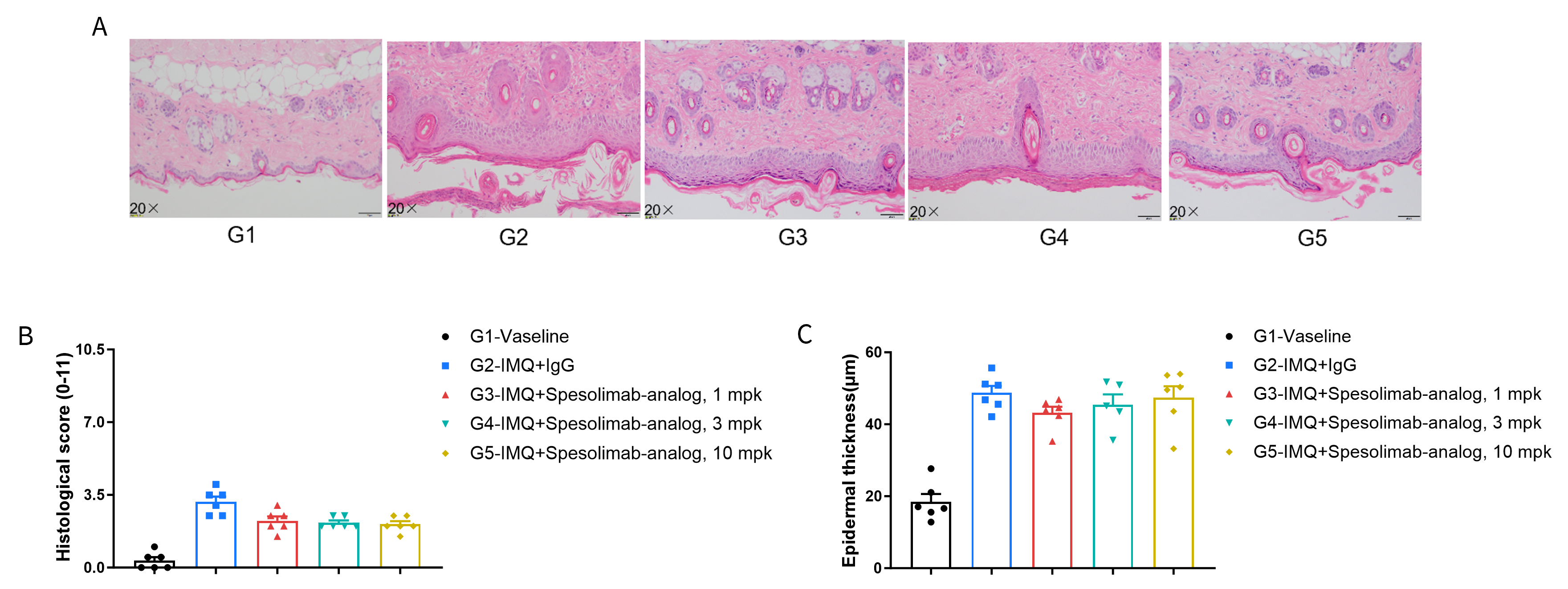
- Spesolimab-analog had therapeutic effects on psoriasis-like skin lesions in B-hIL36R mice, confirming that B-hIL36R mice provide a powerful model for in vivo evaluation of anti-human IL36R antibodies.
Dose dependent effects of antibodies on keratinocyte proliferation and inflammatory cell infiltration in IMQ induced psoriasis-like skin lesions in B-hIL36R mice. Back skin was collected at the endpoint and stained with Hematoxylin and eosin (H&E). (A) H&E staining of the back skin. (B) Histological changes were scored. (C) Epidermal thickness of the mice. Results indicated that Spesolimab-analog (in house) had therapeutic effects on psoriasis-like skin lesions in B-hIL36R mice, confirming that B-hIL36R mice provide a powerful model for in vivo evaluation of anti-human IL36R antibody. Values are expressed as mean ± SEM.
* When publishing results obtained using this animal model, please acknowledge the source as follows: The animal model [B-hIL36R mice] (Cat# 110084) was purchased from Biocytogen.